|
The main task of any sexually reproducing organism is to mate and reproduce. This applies to planktonic copepods as well.
Mating requires meeting, and in the 3-dimensional world of the plankton this represents a major challenge. We have studied how copepods meet, and what the implications of mate finding behaviour is for population dynamics, and are presently trying to figure out how important (if at all) sexual selection is for the evolution of copepod behaviour and life histories.
|
|
Mate finding in copepods
|
Centropages typicus
The females of many copepods produce mating pheromones, and in this case females of Centropages typicus (1 mm) deposit an invisible pheromone trail in her wake. The male can find the trail and sniff down the female. The movie first shows a male tracking down a female, then a 3-dimensional reconstruction of the event (female red, male blue) (Bagøien & Kiørboe 2005) (3-D reconstruction by combining videos from two cameras viewing the event at right angles).
|
|
|
Oithona davisae
The females of the tiny copepod Oithona davisae (0.3 mm) sits still in the water waiting for prey and jumps to a new position every few seconds. This way she produces an interrupted pheromone trail (like a dotted line in the water). Once a male has found one scent patch, he continues circling around this position until he finds the next, and the next,…., and eventually the female. The female tries to escape in a series of jumps, but the male follows her and eventually captures her and completes his mission. A mirror has been placed in the diagonal of the aquarium allowing a view of the copepods as well as of their mirror images, which allows 3-d reconstruction (Kiørboe 2007).
|
|
|
Pseudocalanus elongatus
The females of Pseudocalanus elongatus creates a feeding current that keeps her in a more or less constant position (hovering). The orientation of her body changes all the time, and she thus disperses a cloud of pheromones around herself. A male that has found the pheromone cloud and the female and he now courts her. The significance of the courtship remains unknown (Kiørboe & Bagøien 2005).
|
|
|
Acartia
In some species, mate signalling is hydromechanical rather than chemical. This applies to the species of Acartia in which it has been examined. Mating is preceeded by a series of tandem-hops, which presumably serves the dual purpose of mate detection and identification (Bagøien & Kiørboe 2005).
|
|
Evolution of swimming strategies
In pelagic copepods it is typically the male that has to find the female; thus, the male has to swim, and the faster he swims, the more females he will find. However, (fast) swimming also has costs in terms of energy expenditure and elevated risks of encountering predators. As a consequence, male copepods typically swim faster and more risky than the females. Oithona davisae may provide an example (Kiørboe 2008).
|
Feeding behavior
Both males and females of Oithona davisae are ambush feeders, that is, they remain stationary in the water while waiting for motile prey to pass by. The prey is perceived hydromechanically.
|
|
|
Females hardly move
Females feed most of the time – that is, they hardly move.
|
|
|
Males swin half the time
Males swim about half the time, and when they swim they swim very fast. In fact, male Oithona davisae has a world record in sustained relative swimming velocity.
|
|
|
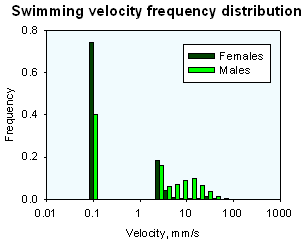
Swimming velocity
Oithona males and females swim at very different velocities. The observed velocities are well predicted by simple models that optimize the number of life time mate encounters.
|
 |
Sex and population dynamics
|
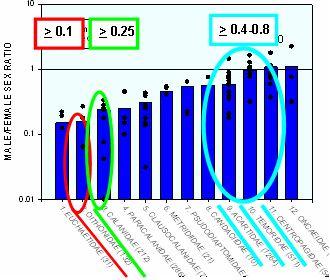
Sex and death
Different male feeding strategies lead to very different swimming strategies. Non-feeding and multi-tasker males that can feed and swim at the same time swim at moderate rates, whereas ambush feeding males swim very fast when they swim. The different strategies lead to very different male mortalities. Since the sex-ratio is close to 1:1 at maturation, gender differences in mortality lead to very different adult sex-ratios. Predictions from simple models are consistent with those observed. Numbers in squares correspond to predicted male:female sex-ratios for different groups of copepods (color-codes).
|
|
|
There are too few males
Because males swim into death in their search for females, there may be too few males to fertilize all females in the population. In the copepod Oithona davisae, the very female-biased sex-ratio implies that only about 1/3 of the females have been fertilized (data from Uye & Sano - Mar Ecol Prog Ser 118: 121-128) (Kiørboe 2007).
|
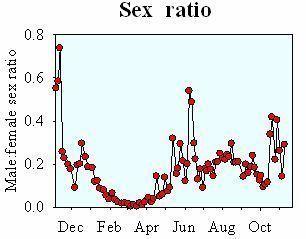
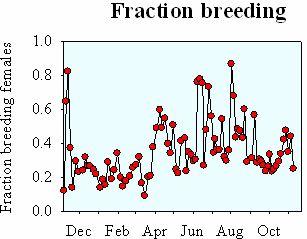
Data from Uye & Sano.
|
|
Allee effect
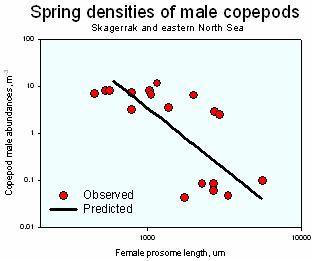
For sexually reproducing organisms, there will be a minimum critical population density below which mate encounters are too rare to allow population maintenance. This density is a function of the mate search capacity (see above), the fecundity, and the mortality. This information combined (in a proper way) allows predicting the seasonal population density of field populations of pelagic copepods. The plot shows observed seasonal minimum densities of adult male copepods in the North Sea and the predicted critical density for the same species (Kiørboe 2006 & erratum)
|
 |
Sexual selection: ongoing work
Do pelagic copepods compete for mates? Do they choose among potential mates? This question of sexual selection is essentially unknown, but a number of observation suggest that this is possible: the earlier maturation of males than of females may suggest a race between males to first find virgin females; the elaborate courtship behaviour found in some species (see movies of Pseudocalanus and Oithona above) may be a way to examine the fitness of the partner; the fertilization limitation and shortage of males found in some species may be suggestive of sexual selection.
Back to list of research areas